|
|
Основные
механизмы генной инженерии Технология рекомбинантной ДНК
Вот мы и добрались до генной инженерии, вызывающей у непосвященных
бурю всевоз-можных эмоций. Если кратко, то суть генной инженерии сводится к следующему:
биологи, зная,
какой ген
за что отвечает, выделяют его из ДНК одного организма и встраивают в ДНК другого.
В результате можно заставить клетку синтезировать новые белки, что придает организму
новые свойства.
Мы знаем, что обмен генетической информацией
происходит и в природе, но только между особями одного вида. Коты «ухаживают» за
кошками, лисы за лисицами, кролики за крольчиха-ми... Случаи же скрещивания особей
разных видов (например, собаки и волка) хоть и случают-ся, но являются скорее исключениями
и возможны лишь для близкородственных животных.
Перенос
генов от родителей к потомкам внутри одного вида называется вертикальным. Так как возникающие при этом особи, как правило, очень похожи на
родителей, в природе генетический аппарат обладает высокой точностью и обеспечивает
постоянство каждого вида.
Генная инженерия дает возможность преодолевать
межвидовые барьеры и передавать признаки одних организмов другим, осуществляя такие
изменения генома, которые вряд ли
могли бы
возникнуть естественным путем. Грубо говоря, генные инженеры делают то, что всегда
запрещала природа. Берут, например, ген из рыбы и вставляют его в помидор. Но не
для того, чтобы помидор плавал, а чтобы его можно было хранить при низкой температуре.
Перенос генов между особями разных видов называется горизонтальным (латеральным).
Молекула ДНК, собранная из кусочков ДНК различных
организмов, называется рекомби-нантной. Первая
рекомбинантная ДНК, объединившая гены обезьяньего вируса SV40,
бактериофага
лямбда и галактозного оперона E.coli, была создана в 1972 году группой
америка-нских ученых под руководством П. Берга. Однако первые “генные операции”
такого рода эффективно проводились уже более 4,5 миллиардов лет назад главным “генным
инженером” – Природой. Речь идет об уже знакомых нам прокариотах – первых живых
существах, населявших нашу планету еще со времен Архейской эры. Как уже говорилось,
механизмы передачи генетической информации в природе обладают высокой стабильностью,
призванной служить сохранению и выживанию вида. Стабильность эта, однако, не абсолютна,
ведь иначе в ходе эволюции не смогло бы возникнуть такого многообразия форм жизни,
свидетелями (и представителями) которых мы являемся.
Но прокариоты, как известно, размножаются простым
делением, при котором каждая дочерняя клетка получает «в наследство» точную копию
родительской ДНК. Откуда же возникло такое разнообразие в среде самих простейших
и как могли появиться другие организмы? Одна из наиболее очевидных причин генетической
изменчивости – это мутации, являющиеся, по меткому выражению Дарвина, двигателем
эволюции.
Мутации – cкачкообразные изменения генетического кода клетки, приводящие
к появлению новых признаков. Если потомки измененной особи имеют некоторое преимущество
перед обычными, например, большую жизнеспособность или повышенную скорость роста,
они постепенно накапливаются и вытесняют исходных особей. Различают мутации спонтанные (причины их возникновения
неизвестны) и индуцированные. Индуцировать
мутации могут различные факторы, действующие на генетический материал клетки: физические,
химические или биологические.
В ходе эволюции прокариоты выработали способы
защиты своего генетического материала от повреждающего действия облучения, химических
веществ и других мутагенов. В их клетках обнаружены эффективные системы ремонта
поврежденных участков ДНК. Если бы таких механизмов не было, то организм бы переродился
и вымер как вид. Основной механизм вос-становления ДНК – это “вырезание” повреждений,
так называемая рестрикция. Ее осуществля-ют
ферменты эндонуклеазы, расщепляющие нить ДНК. Такой способ помогает, только если повреждена
только одна цепочка молекулы. Тогда поврежденный участок вырезается, а образо-вавшаяся
брешь заполняется комплементарными нуклеотидами с использованием в качестве матрицы-шаблона
неповрежденной нити ДНК. Такимобразом, многие случайные мутации
попросту
вырезаются.
Однако когда повреждение касается обеих нитей,
тот же самый механизм восстановления превращается в орудие самоубийства: эндонуклеазы
распознают поврежденный участок и разрывают в его месте обе нити ДНК. Кстати говоря,
то же самое происходит и в клетках многоклеточных организмов в случае фатального
повреждения хромосом. Такое генетически запрограммированное самоубийство биологи
называют апоптозом. Оно сохраняет
«чистоту генов» и предохраняет вид от деградации. Можно сказать, что клетка руководствуется
своеобразным “клеточным самурайским законом” – “лучше умереть, чем ошибиться!”
Однако присущий всем живых существам инстинкт
самосохранения порой все же берет верх, и клетке_мутанту удается “обмануть” убийственную
рестрикцию путем модификации ДНК
метилированием
или введением дополнительных пар нуклеотидов. Уцелевшая клетка приобретает новые
свойства и, если они оказываются выгодными, дает начало новому виду существ.Итак,
огромное разнообразие организмов объясняется мутациями. Но, как оказалось, тому
есть и другие причины. С развитием генетики ученые обнаружили, что для прокариот
ха-рактерен путь горизонтального переноса генов между различными особями.
Молекулярно_генетический анализ показал, что
геномы прокариот представляют собой мозаику генов, приобретенных у разных видов.
Одинаковые генетические последовательности можно увидеть у многих прокариот, вне
зависимости от степени их родства. Объяснить воз-никновение такой мозаики может
только горизонтальный перенос генов.
Горизонтальный перенос
генов у прокариот – это не просто лабораторный арте-факт или результат генной инженерии,
а распространенное природное явление.
Установлены три основных механизма латерального
переноса: трансформация, коньюгация и трансдукция.
Трансформация – это нормальная физиологическая функция
обмена генетическим материалом у некоторых бактерий.
Конъюгация имеет наименьшее число ограничений
для межвидового обмена генетической информацией, но предполагает тесный физический
контакт между микроорганизмами,легче всего достижимый в биопленках.
Трансдукция (от лат. transductio – перемещение)
– это перенос генетического материала из одной клетки в другую с помощью некоторых
вирусов (бактериофагов), что приводит к измене-нию наследственных свойств клетки_реципиента.
Явление трансдукции было открыто американскими учеными Д. Ледербергом и Н. Циндером
в 1952. Последний механизм следует рассмотреть более подробно,но прежде скажем несколько
слов о вирусах.
Вирусы (от лат. virus – яд) были открыты в
1892 г. русским ученым_ботаником Д. И. Ивановским при изучении мозаичной болезни
(пятнистости листьев) табака. К наиболее опасным заболе-ваниям, вызываемым вирусами
у животных и человека, относят бешенство, оспу, грипп, полиомиелит, СПИД, гепатит
и др.Вирусы – существа совершенно удивительные. Они занимают промежуточное положение
между живой и неживой материей, представляя собой случай некого биологического дуализма.
На вопрос “живые ли вирусы?” нельзя ответить однозначно, ведь если живой считать
структуру, способную к размножению и обладающую наследственной информацией, то можно
сказать, что вирусы живые (впрочем, по этому определению живыми можно признать и
компьютерные вирусы). Но если считать живой структуру, обладающую клеточным строением
(как, например, растения, грибы, животные), то ответ должен быть отрицательным.
Следует также отметить, что вирусы не способны воспроизводить себя вне клетки_хозяина.
Вирусы
устроены очень просто. Они состоят из головки (сердцевины) округлой, гексагональной
или палочковидной формы диаметром 45_140 нм и отростка толщиной 10_40 и длиной
100_200
нм. В
1 мм3 воды может уместиться около миллиарда фагов.
|
|
|

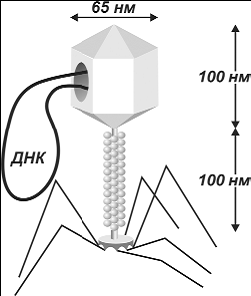
Рис 187. Электронно_микроскопическая фотография и схема строения
бактериофага T4
Сердцевина вируса содержит ДНК или РНК. Ее
окружает защитная белковая оболочка (капсид). Отросток имеет вид полой трубки, окруженной сократительными белками вроде мы-шечных. На конце отростка
у многих вирусов есть базальная пластинка, от которой отходят
тонкие длинные отростки_«ноги», служащие
для прикрепления к клетке_хозяину. Вирус может воспроизводить себе подобных только
внедряясь в клетку хозяина. Для этого он прикрепляется к бактериальной клетке и
растворяет клеточную стенку, выделяя особый фермент. Затем содержимое головки по
канальцу отростка проникает в клетку. Попав внутрь клетки, вирус, подобно своему
компьютерному собрату, перепрограммирует рибосомы на производство своих копий. Он
«выключает» хозяйскую ДНК и, используя свою собственную ДНК или РНК, «заставляет»
клетку синтезировать новые копии вируса. После сборки большого числа вирусных частиц
клетка, как правило, погибает, а множество новых вирусов, произведенных с ее по-мощью,
выходит на свободу, поражая другие клетки. Вирусы, вызывающие гибель клетки, называют вирулентными19.
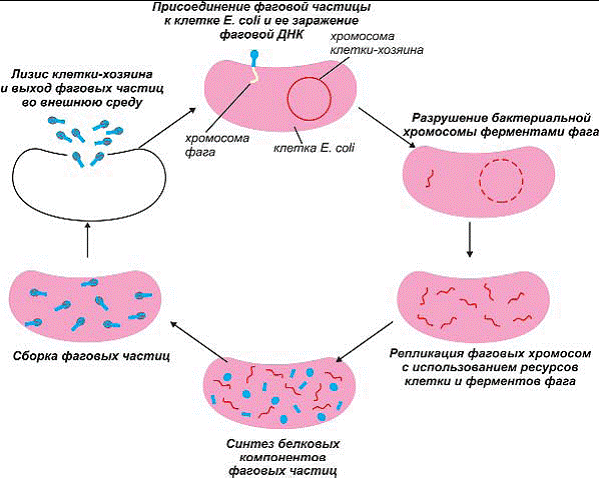
Рис 188. Жизненный цикл вирулентного фага, например Т2 или Т4
19 Вирулентность – это степень болезнетворного действия микроба.
Ее можно рассматривать как способность адаптироваться к организму хозяина и преодолевать
его защитные механизмы.
Однако существует и другая категория вирусов,
называемых умеренными(симбиотически-ми). Проникая внутрь
клетки,они могут либо повести себя как обычные фаги, вызывая гибель клетки, либо
могут встроиться в ДНК клетки_хозяина и остаться в ней в скрытой неинфекцион-ной
форме.
Клетки, содержащие такой вирус, называются
лизогенными, они могут содержать 2, 3 и более фагов. Лизогения может затем передаваться
потомству бактерии. На рисунке показаны два возможных пути развития умеренного вируса.
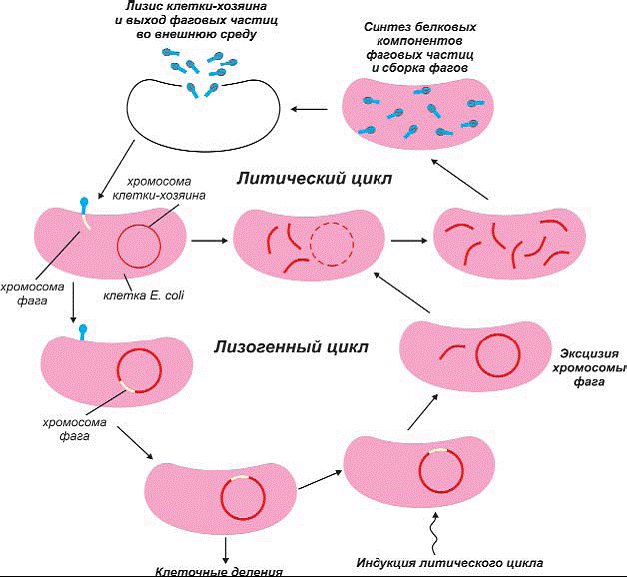
Рис 189. Жизненный цикл умеренного фага. Когда он инфицирует
клетку, развитие может пойти
по литическому или лизогенному пути
Так, а что же наш «горизонтальный перенос»?
Напоминаем, что трансдукция – это перенос вирусом бактериальных генов из одной клетки
в другую, что приводит к изменению наследствен-ных свойств клетки-реципиента.
Трансдукция возможна, если в процессе размножения умеренного
фага одна из частиц вирусной ДНК случайно захватит фрагмент бактериальной хромосомы
Когда впоследствии такой вирус заражает другую
бактерию, участок бактериальной ДНК проникает в клетку таким же путем, как вирусный.
Между трансдуцированной ДНК и участком хромосомы может произойти обмен, и как следствие
его возникают рекомбинанты, несущие небольшую
часть генетического материала клетки-донора.
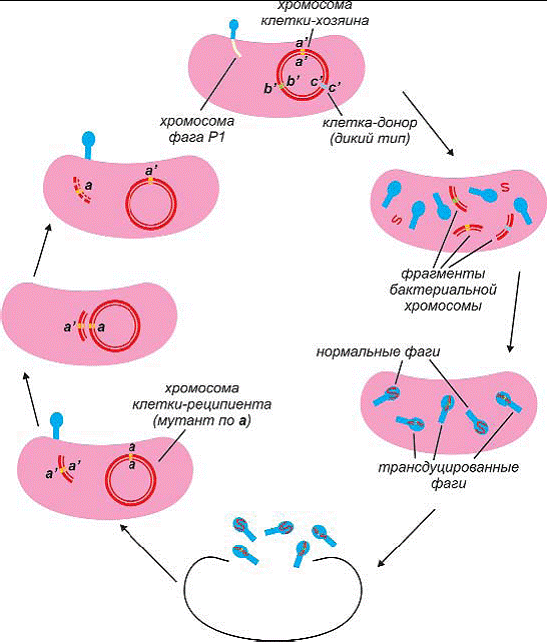
Рис 190. Схема общей трансдукции между линиями Е. coli:
а) Клетка дикого типа, инфицированная фагом P1;
б) ДНК клетки_хозяина деградирует в ходе литического цикла;
в) в ходе сборки фаговых частиц некоторые фрагменты бактериальной
хромосомы
включаются в несколько фагов_потомков, что потом приведет к
трансдукции;
г) лизис;
д) трансдуцирующий фаг инфицирует бактерию_реципиента;
е) происходит обмен донорного гена а+ и реципиентного гена а_;
ж) образование стабильного трансдуктанта а+.
Итак, мы рассмотрели процесс естественного
“горизонтального” обмена генетической информацией между бактериями, то есть перемещения
генов из одного организма в другой посредством мобильных генетических посредников. Рассмотренный механизм
получил назва-ние трансдукции, а понимание его сути привело к рождению генной инженерии.
Как уже было сказано, суть генной инженерии заключается в том, что в генотип организма
встраиваются или исключаются отдельные гены или их группы. Для этого используют
метод получения рекомби-нантных, т. е. содержащих чужеродный ген, ДНК, которые затем
вводятся в организм реципиента и становятся составной частью его генетического аппарата.
Этот процесс состоит из нескольких этапов:
1. Чтобы
выделить тот или иной ген из цепочки ДНК, используется рестрикция – разрезание ДНК на фрагменты уже знакомыми
нам ферментами-рестриктазами. Они попросту «шинкуют» ДНК: режут на отрезки, но не
как попало, а в определенных местах. Всякая рестриктаза может опознать лишь одну
стандартную последовательность из нескольких нуклеотидов. Молекулы рестриктазы химически
связываются с ними и в этих местах рвут цепь ДНК. На рисунке изображена схема расщепления
ДНК по остаткам А (аденин).
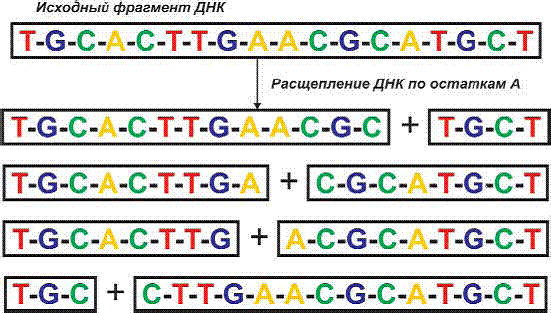
Рис 191. Схема получения фрагментов ДНК в результате расщепления
по
определенному нуклеотиду (А)
В настоящее время известно более 400 рестриктаз,
способных расщеплять ДНК по 120 различным последовательностям нуклеотидов.
2. Лигирование – процесс «сшивания» генов с помощью
особых ферментов, называемых лигазами. Лигазы сшивают участки ДНК, образовывая между
их крайними нуклеотидами
химическую
связь.
3. После
того как рекомбинантная ДНК сшита, ее вводят в живые клетки. Этот процесс называется
трансформацией. Для того, чтобы рекомбинантная ДНК стала частью генетического
аппарата
клетки, она должна либо встроиться в ее геном и реплицироваться за ее счет, либо
быть способной к автономной репликации. Для этого используют векторы – мобильные гене_
тические
элементы: вирусы, плазмиды и транспозоны. Эти элементы могут присоединять те или
иные гены к своей ДНК, а затем, оказавшись в клетке_хозяине, встраиваться вместе
с «оторван-ным» чужеродным геном в хромосому хозяина, которая потом реплицируется
уже вместе со всей этой новой последовательностью. В общих чертах это напоминает
трансдукцию, имеющую, как мы убедились, место и в природе.
Итак, мы познакомились с общими принципами
искусственного конструирования молекул ДНК. Рассмотрим теперь несколько примеров
практического применения генной инженерии.
|
|